Exogyra plexa is a small, approximately 1 cm sized, oyster that occurs in the Fredricksburg and Washita formations of the Cretaceous deposits of Texas. It is thought to have evolved from a Ceratostreon ancestor. It is also thought to be the ancestor of Ilymatogyra arietina and to have given rise to a major radiation of Exogyrid oysters beginning in the Cenomanian. Here collections of E. plexa, from the Goodland, Kiamichi, and Duck Creek Formations, and I. arietina, from the Main Street and Grayson Formations, are described. These collections indicate some of the processes by which E. plexa may have been transformed into I. arietina. In particular, the upper valves show a nice grade, going from the oldest E. plexa to the youngest I. arietina, in the reduction of the area of the concave portion of the shell. The lower valves indicate several, although somewhat more erratic, trends. These include a trend towards reduced attachment scars, increase in beak height, and reduction of undulations and shell ribbing. Also, a Ceratostreon texanum specimen from the same locality at which the E. plexa of the Goodland Formation were collected is described in order to point out some of the features that indicate Ceratostreon ancestry for E. plexa.
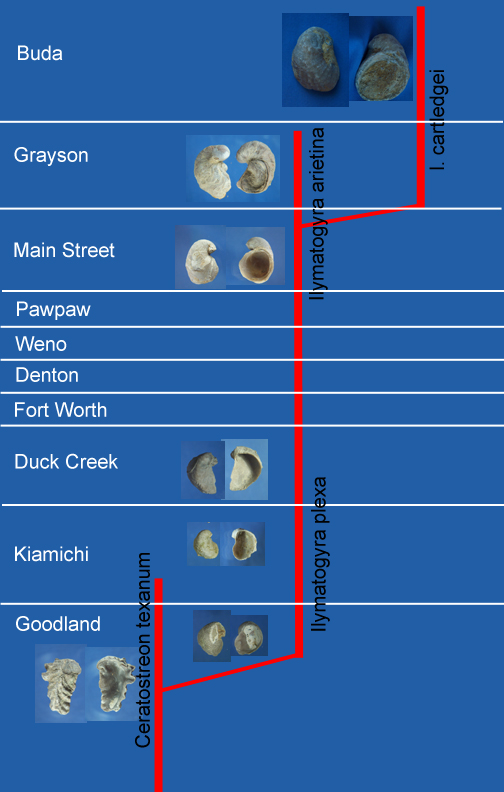
Figure 1 depicts a phylogeny of Exogyrid oysters proposed by Malchus in 1990. It shows the E. plexa lineage branching off of the Ceratostreon lineage and then giving rise to a major radiation of Exogyrid oysters. It is useful to remember that such a phylogeny is a theory or hypothesis of how organisms are related through descent. As with any good scientific theory, such phylogenies stand or fall, depending on how well they hold up when subjected to new data. In the present case, new data is supplied through the discovery of new fossils or through the measurement of new characters of old fossils. In recent years, the study of shell microstructure has supplied new characters to test fossil oyster phylogenies. Malchus, among others, has contributed significantly to our knowledge of shell microstructure [1], and the phylogeny depicted in Fig. 1 incorporated this knowledge. Nevertheless, a few more things have been learned about fossil oysters since 1990, so we would draw a somewhat modified phylogeny today. The topic treated here, however, is that of the Ceratostreon-Exogyra plexa-Ilymatogyra arietina lineage, one of the more secure parts of this phylogenetic tree. Malchus had observed that both E. plexa and I. arietina are North American species and he suggested that a careful examination of the Cretaceous strata of North America might reveal transitional forms between these two species. This peaked the interest of Charles and me in examining E. plexa and I. arietina populations at various stratographic levels.
|
| Figure 1. A phylogeny of Exogyrid oysters proposed by Malchus [1] in 1990. The range of occurrence of each oyster species is indicated by a thick vertical red bar. The position of occurrence within the geologic column is referenced to the European stages indicated in the column on the left-hand side of the figure. |
We were further intrigued by a paper written by Böse [2] in 1919. Figure 2 shows the phylogeny of Exogyrid oysters suggested in that paper. Here the figure has been somewhat modified from that given in the Böse paper. In particular, we have updated the names of the formations and the genus and species names to correspond with current usage. Following the lineage on the right-hand side of this phylogeny, it is evident that Böse thought that populations of E. plexa and I. arietina occurring at different stratographic levels were sufficiently distinguishable that some of them should be regarded as new species. This suggested to us that Böse, already some 70 years earlier, may have found some of the intermediate forms that Malchus had predicted in 1990 could be found by careful examination of North American strata. To explore this possibility, Charles and I have made reference collections of populations of E. plexa and I. arietina from stratographic levels similar to those examined by Böse. Here these collections are described and we discuss their implications for Exogyrid phylogeny.

|
| Figure 2. A phylogeny of Texas Exogyrid oysters proposed by Böse [2] in 1919. |
| Figure 3. Top view of an E. plexa upper valve with the convex and concave regions indicated. This specimen is from the Goodland Formation. The length of the valve is 1.31 cm and the width is 1.04cm. |
The collections of E. plexa and Ilymatogyra arietina upper valves from successively higher strata show a graded trend where the area of the concave region is reduced relative to that of the convex region. This trend is illustrated in Figure 4, where specimens have been selected to indicate the range of variability of the upper valves in successive formations. The extremes of the variation in successive populations overlap as can be seen by comparing the E. plexa valve, 4a, from the Goodland Formation, with the E. plexa valve, 3b, from the Kiamichi Formation, and the E. plexa valve, 3a, from the Kiamichi Formation with the I. arietina valve, 2b, from the Main Street Formation. Note that the I. arietina valve, 2a, from the Main Street Formation, is very similar to the upper valves, 1a and 1b, of I. arietina, typical of the Grayson Formation.

|
| Figure 4. Top view of upper valves of the E. plexa and I. arietina, showing the trend in which the area of the concave region is reduced while the area of the convex region increases. 1a and 1b are I. arietina from the Grayson Formation. 2a and 2b are I. arietina from the Main Street Formation. 3a and 3b are E. plexa from the Kiamichi Formation. 4a and 4b are from the Goodland Formation. For easy size comparison, all the images are set to the same linear scale. Valve 4a is the same as that of figure 3, where the dimensions are given. |
As the convex region of the upper valve becomes larger and the concave region is reduced, the valves become more dome-shaped. This is illustrated by the side views displayed in Figure 5.

|
| Figure 5. Side views of upper valves of the E. plexa stalk. 1a and 1b are different side views of the same upper valve of an I. arietina from the Grayson Formation. 2a and 2b are different views of the same valve of an I. arietina from the Main Street Formation. 3a and 3b are different views of the same valve of an E. plexa from the Kiamichi Formation. 4a and 4b are side views of the same valve of an E. plexa from the Goodland Formation. The "a" views are along the short axis of the valve. The "b" views are along the long axis of the valve from the direction of the beak. For easy size comparison, all the images are set to the same linear scale. The length of valve 1a is 2.41 cm. |

|
| Figure 6. Side view of lower valve; view from posterior end. Valve 1 is that of an I. arietina from the Grayson Formation. 2a and 2b are I. arietina valves from the Main Street Formation. Valve 3 is that of an E. plexa from the Duck Creek Formation. 4a and 4b are E. plexa valves from the Kiamichi Formation. Valve 5 is that of an E. plexa from the Goodland Formation. All images are set to the same linear scale for easy size comparison. The largest dimension shown of the valve in image 1 is 3 cm. |
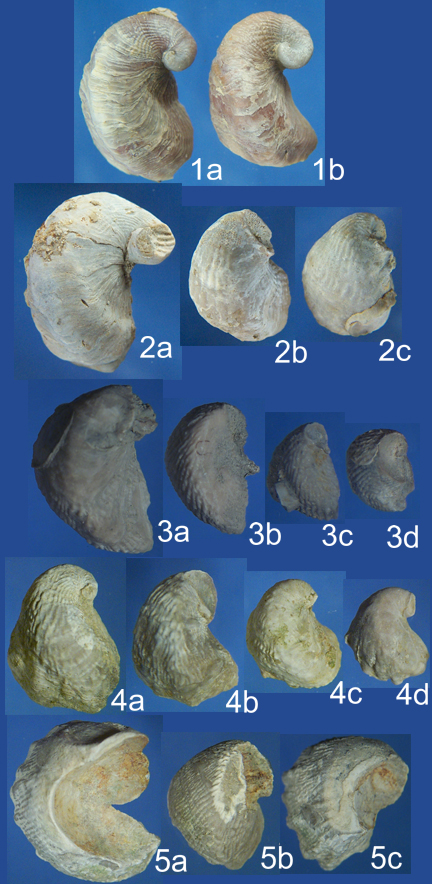
Lower views of the lower valves shown in Figure 7 illustrate further trends in the evolution of the E. plexa and I. arietina stalk. The trend toward reduction of the folds and the ribs is evident. The I. arietina of the Grayson Formation only possesses ribs near the tip of the beak. Also, there is a trend toward a reduction of the attachment area. For specimens of E. plexa from the Goodland Formation, such as 5a, the attachment scar is often comparable to the valve itself in size. For I. arietina of the Grayson Formation an attachment scar is generally absent.
| Figure 7. Lower view of lower valves. 1a and 1b are I. arietina from the Grayson Formation. 2a-2c are I. arietina from the Main Street Formation. 3a-3d are E. plexa from the Duck Creek Formation. 4a-4d are E. plexa from the Kiamichi Formation. 5a-5c are E. plexa from the Goodland Formation. All images are set to the same linear scale for easy size comparison. The height of the image 1a is 2.28 cm. |
Upper views of the lower valves are shown in Figure 8. The end members of this sequence, that is, the Exogyra plexa from the Goodland Formation and the Ilymatogyra arietina from the Grayson Formation, possess upper valves whose shell is somewhat thicker than that of the specimens from the intermediate formations. As indicated in Figure 1a, the I. arietina of the Grayson Formation often show a well-developed spiral concavity that is rarely present in the I. arietina of the Main Street Formation.

|
| Figure 8. Upper view of lower valve. 1a and 1b are different views of the same I. arietina specimen from the Grayson Formation. This specimen is also depicted in image 1 of Figure 6. In figure 1a the spiral concavity is pointed out. 2a and 2b are I. arietina from the Main Street Formation. 3a through 3c are from the Duck Creek Formation. 4a through 4d are from the Kiamichi Formation. 5a through 5c are from the Goodland Formation. All images are set to the same linear scale for easy size comparison. The height of the image of valve 1a is 3.9 cm. 1a and 1b are different views of the same shell. |

|
| Figure 9. Ceratostreon texanum from the Goodland Formation, lower view of lower valve. The length of the valve is 2.8 cm; the width is 1.83 cm. |

|
| Figure 10. Ceratostreon texanum, upper view of the same lower valve shown in Figure 9. |

|
| Figure 11. Ceratostreon texanum, view of beak area of the same lower valve shown in Figures 9 and 10. The width of the valve is 1.83 cm. |

|
| Figure 12. Views of the beak area of the lower valve of two Exogyra plexa specimens from the Goodland Formation. Both images are set to the same linear scale. Other views of the valve "a" can be seen in image 1 of Figure 13, and by clicking on the image in Figure 19 located in the Lower Valve column and the Goodland row. Views of this valve are located in entries 1a, 1b, 7, and 8 of the resulting table of clickable images. Valve "b" is also depicted in Figure 5c. |
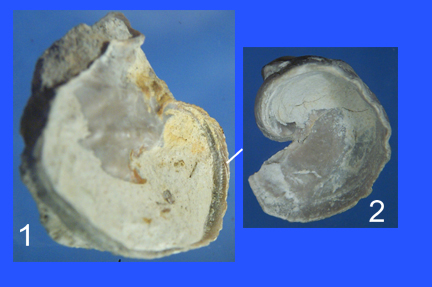
As can be seen by a perusal of the images that can be obtained by clicking on the images in Figure 19, the reference collection indicates an evolutionary trend towards a more stereotypical lower valve shape. The E. plexa of the Goodland Formation are quite variable. The degree of variability decreases in younger rock formations. The variability I. arietina lower valves of the Grayson Formation is remarkably small for oysters. Figure 13 shows two extreme examples of E. plexa lower valves from the Goodland Formation that suggest descent from a Ceratostreon ancestor. Both valves possess large attachment scars, and the valves themselves are larger than typical. The valve in image 1 of Figure 13 exhibits chomata along a large fraction of the shell margin as is typical of Ceratostreons. Such chomata can be seen along the shell margin of the C. texanum depicted in Figure 10. More typically, in E. plexa and I. arietina chomata are limited to an area near the beak. Examples of this can be seen by close inspection of images that can be obtained by clicking on the images in the first column of Figure 19. The E. plexa lower valve depicted in image 2 of Figure 13 exhibits thickening of the shell wall in the beak area, reminiscent of the growth habit of Ceratostreons. The high degree of variability of the E. plexa of the Goodland Formation, their tendency toward forms resembling Ceratostreons, suggests that the split of the E. plexa from the Ceratostreons occurred during the time the Fredricksburg strata, to which the Goodland Formation belongs, were deposited. Support for this also comes from the apparent absence of E. plexa from older strata of the Fredricksburg Division, namely, the members of the Walnut Marl, which are nevertheless highly fossiliferous.
|
| Figure 13. Upper view of the lower valve of two Exogyra plexa specimens from the Goodland Formation. Valve 2 is also depicted in images 5a of Figures 7 and 8. Views of this valve can also be accessed by clicking on the image in the Lower valve column and Goodland row of the image table in Figure 19. This valve is depicted in entries 2a, 2b, and 9 of the resulting image table. The height of the image of valve 2 is 2 cm. |
Figure 14 presents three views of a lower valve which I have difficulty in deciding to assign to C. texanum or E. plexa. It possesses an attachment scar that covers most of the shell surface as can be seen in "b." The shell does not exhibit the folds that C. texanum typically displays. It is quite different from the more typical specimen of Figures 9, 10, and 11, for example. It shows ribbing typical of E. plexa, but only faintly. Clearly, it would be informative to build a collection documenting the degree of variability C. texanum exhibits in the Goodland Formation.

|
| Figure 14. Is it an E. plexa or a C. texanum? The length of this valve is 2.7 cm. |
 1a 1a
|
 2a 2a
|
 2c 2c
|
 3a 3a
|
 1b 1b
|
 2b 2b
|
 2d 2d
|
 3b 3b
|
| - | |||
 4a 4a
|
 5a 5a
|
 5c 5c
|
 6a 6a
|
 4b 4b
|
 5b 5b
|
 5d 5d
|
 6b 6b
|
| - | |||
 6c 6c
|
 6d 6d
|
||
| Figure 15. Exogyra cartledgei. Click on images for a larger view. All the images are set to the same linear scale. The width of each image is 6.3 cm. | |||
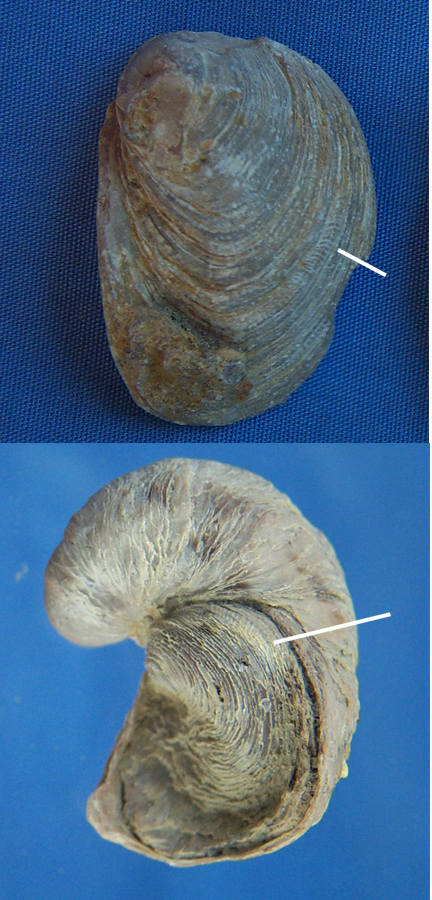
Image 1 of Figure 16 is a side view of the upper valve of E. cartledgei. For comparison, using the same size scale, upper valves of I. arietina and E. plexa are also shown in the figure. The white lines point out a groove that develops along the rim of the E. cartledgei and I. arietina valves, due to the way shell material is added to accommodate the twist as the oyster grows. Although much less developed, some E. plexa upper valves also display this kind of growth pattern. This feature supports an Ilymatogyra ancestry for E. cartledgei.

|
| Figure 16. Upper valves, anterior view. 1, E. cartledgei from the Buda Formation. 2a and 2b, I arietina from the Grayson Formation. 3a and 3b are I arietina from the Main Street Formation. 4 and 5 are E. plexa specimens from the Kiamichi and Goodland Formations, respectively. Images are all set to the same linear scale for easy size comparison. The width of the image of valve 1 is 4.6 cm. |
|
| Figure 17. Views of an upper valve of Exogyra cartledgei (upper image) and an articulated specimen of Ilymatogyra arietina (lower image). Length of valve in upper image is 3.10 cm. Height of the image of the specimen in the lower image is 2.75 cm. |
Böse proposed the existence of two distinct species of Exogyra plexa-like oysters in strata below the Kiamichi Formation [1]. See Fig. 2. The distinction between the two species was based on whether the oyster "developed on the anterior side four to five strong radial folds beginning near the umbonal ridge and going down obliquely with the ridge to the antero-inferior margin." Since he did not have enough material to allow a complete description of the species, Böse designated it as Ex. nov. sp. aff. plexa. Our own collection of material from the Goodland Formation does not seem to support such a distinction. The radial folds occur with a range of amplitudes that does not allow a natural division of the specimens into two distinct groups. Instead, E. plexa appears to have been quite variable during the time the Goodland limestone was deposited. The radial folds can be interpreted as having been inherited from C. texanum, as species in whic h the folds are also highly variable.
To Böse the E. plexa-like oysters from the Kiamichi Formation and the base of the Fort Worth Formation were sufficiently different that each represented a different species or variety. The form found in the Kiamichi was described as showing "an ornamentation very similar to that of Ex. plexa on the umbonal ridge on the posterior side and several obtuse folds covered with fine radial ribs on the anterior side near the antero-inferior margin." In our collection from the Kiamichi there are specimens that fit this description, although the form of individual specimens can deviate significantly from this description. In addition to the distinction Böse noted, we would add that the shell thickness of both valves in our collections are generally thinner than those of the Goodland Formation. In addition, the convex area of the upper valve covers a larger portion of the valve surface.
Böse comments that "Ex. plexa seems to lose the fine ribs on the lower margin in larger specimens ... This feature is still more evident in larger specimens found at the base of the Fort Worth limestone ... It is evidently a younger form of the same group, but possibly a different species or variety. It grows much larger than the type, shows approximately the same ornamentation on the umbonal region, but becomes entirely smooth on the inferior margin." Our specimens from the Duck Creek Formation do not exhibit this ornamentation pattern, perhaps because the specimens are not of sufficiently mature individuals. Our specimens were collected from the Lower Duck Creek Formation near the boundary with the Kiamichi Formation. So, alternatively, it may be that the E. plexa of the Fort Worth Formation are significantly different from those of the lower Duck Creek Formation.
| Figure 18. The phylogeny of the Exogyra plexa stalk proposed here. |
| Formation | Lower valve | Front view | Upper valve | Side view |
| Grayson |

|

|

|

|
| Main Street |

|

|

|

|
| Duck Creek |

|

|
||
| Kiamichi |

|

|

|
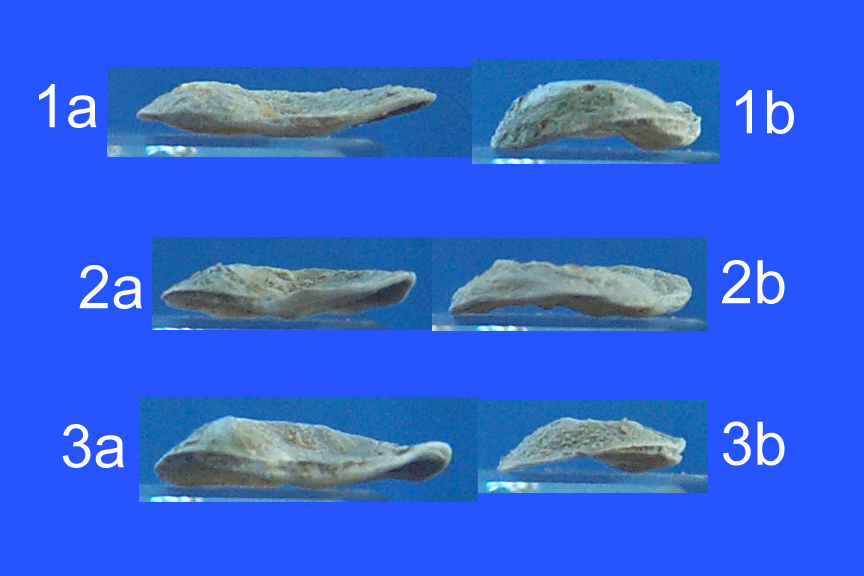
|
| Goodland |

|

|

|

|
| Figure 19. Images of Exogyra plexa and Ilymatogyra arietina specimens in my reference collection. Click on image to bring up a further set of images which can be clicked on to obtain an enlarged view of the image. All images accessed in the Lower valve column are 5.3 cm in width. All images accessed through the Front view and Side view columns are 6.6 cm in width. All images accessed through the Upper valve column are 3.5 cm in width. | ||||
[1] N. Malchus, Revision der Kreide-Austern (Bivalvia: Pteriomorphia) Ägyptens (Biostratigraphie, Systematik), Berliner geowiss. (A) 125, (1990), page 194.
[2] E. Böse, "On a New Exogyra from the Del Rio Clay and Some Observations on the Evolution of Exogyra in the Texas Cretaceous," Bulletin No. 1902 of the University of Texas Bureau of Economic Geology, (1919).
[3] E. A. Mancini, "Origin of the Grayson Micromorph Fauna (Upper Cretaceous) of North-Central Texas," Journal of Paleontology 52 1294-1314 (1978).